
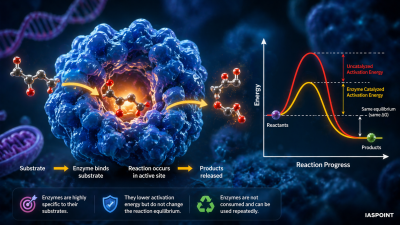
Enzymes function as biological catalysts by providing a specific environment that facilitates chemical reactions. They do not change the equilibrium of a reaction or the nature of the products; instead, they significantly accelerate the rate by reducing the energy barrier required for the reaction to occur.
The Concept of Activation Energy
All chemical reactions require an initial input of energy to reach a high-energy “transition state” before converting into products. This energy is known as Activation Energy (Ea).
- Lowering the Barrier: Enzymes lower the activation energy, allowing a higher proportion of substrate molecules to reach the transition state even at the relatively low temperatures of living organisms.
- Transition State: This is a transient, unstable structural state formed when the substrate binds to the enzyme. It is the peak of the energy profile in a chemical reaction.
Stages of the Catalytic Cycle
The mechanism of enzyme action is a sequential process involving the formation of an enzyme-substrate complex. The steps are as follows:
- Substrate Binding: The substrate molecule diffuses into the Active Site of the enzyme.
- Induced Fit: The binding of the substrate induces a conformational change in the enzyme, causing the active site to wrap more tightly around the substrate.
- Formation of ES Complex: A temporary, non-covalent bond forms between the enzyme and substrate, creating the Enzyme-Substrate (ES) Complex.
- Transition State Formation: The enzyme stresses specific bonds in the substrate, facilitating the conversion of the substrate into products. This results in the Enzyme-Product (EP) Complex.
- Product Release: The enzyme releases the finished products. Because the enzyme remains chemically unchanged, it is free to bind another substrate molecule and repeat the cycle.
Theories of Enzyme-Substrate Interaction
Biochemists have proposed two primary models to explain how enzymes recognize and bind their specific substrates.
1. Lock and Key Hypothesis
Proposed by Emil Fischer, this model suggests that the enzyme’s active site has a rigid, pre-defined shape that exactly matches the substrate, much like a specific key fits a specific lock. While it explains specificity, it does not account for the flexibility seen in most enzymes.
2. Induced Fit Theory
Proposed by Daniel Koshland, this is the more widely accepted modern view. It suggests that the active site is flexible. When the substrate begins to bind, the enzyme undergoes a structural shift to create a complementary fit. This “hand-in-glove” interaction optimizes the catalytic environment.
Factors Determining Catalytic Efficiency
The efficiency of the enzyme mechanism is measured by two primary parameters:
- Michaelis Constant (Km): The substrate concentration at which the reaction rate is half of the maximum velocity (Vmax). A low Km indicates high affinity between the enzyme and its substrate.
- Turnover Number (kcat): The number of substrate molecules converted to product by a single enzyme molecule per second.
Catalytic Strategies Employed by Enzymes
Enzymes use several chemical strategies within the active site to speed up reactions:
- Proximity and Orientation: Enzymes bring reactants close together and align them in the perfect orientation for a collision to result in a reaction.
- Acid-Base Catalysis: Side chains of amino acids (like Histidine) act as proton donors or acceptors to stabilize charges.
- Covalent Catalysis: A temporary covalent bond is formed between the enzyme and the substrate during the transition state.
- Metal Ion Catalysis: Co-factors like Zn2+, Mg2+, or Fe2+ help orient the substrate or stabilize negative charges.
UPSC Prelims Fact File
- Reversibility: Most enzymatic reactions are theoretically reversible; the enzyme catalyzes the reaction in both the forward and backward directions depending on the concentration of reactants and products.
- Specificity: Enzymes are highly specific due to the unique shape and chemical environment of the active site (e.g., Urease only acts on Urea).
- Speed: Carbonic Anhydrase can catalyze the formation of 600,000 molecules of carbonic acid per second, compared to about 200 molecules per hour without the enzyme.
- Thermolability: Because the mechanism relies on a specific 3D protein structure, the mechanism fails at high temperatures (usually above 45°C) due to denaturation.